

My research has had good exposure, being published in 23 different international journals, book chapters, and a monograph, in chemistry, physics, plant physiology, ecology, global change, agronomy, remote sensing, and other fields (evidence of a poor attention span for a given discipline?). Still, there are findings that I wish to have more utility in research and in applications. One example with practical and policy implications is my the formulation that my wife, Dr. Lou Ellen Kay, and I developed of functional balance in plants’ nutrient acquisition and nutrient use (for photosynthesis), particularly as it offers a physiological framework for understanding the drop in tissue N content at elevated CO2. The common explanation for the drop is dilution by high carbohydate accumulation, which is not so useful. I’ve elaborated more on high-CO2 effects, projecting diversity in plant responses in tissue N content, N-use efficiency, water-use efficiency, and growth rate. In climate-change research, many models of diverse (or, alternatively, in-common) responses of plants focus on temperature and precipitation changes, omitting the direct physiological responses to CO2. Another example is my finding, on the modeling side of a modeling-experiment study, that pecan trees apparently lack a water-sparing response to low relative humidity. This has implications for irrigation water use, orchard siting, and other policies.
These under-utilized ideas will be populated on this page and its linked pages. In this effort, especially, I will welcome comments and collaborations. I am still setting up the structure of comment handling at this time.
Three pages are linked from here:
- My published research. You may deem this as research vetted by the peer-review process, for what value that adds.
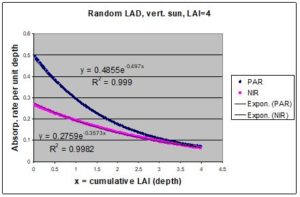
- Here’s some research that recently (Dec. 2017) “leaked” into teaching, as well – a lot about light scattering and absorption
- Ideas for development. Here you will find ideas that I stopped pushing through peer review but that I offer as valuable. An example is my finding that mass flow of water in soil does not increase net delivery of nutrients to plant roots over the rate provided by diffusion in quiescent soil water (flattening of the diffusion gradient cancels the gain from mass flow). Other ideas have not yet been put out for publication. Some are relatively small in scope but intriguing, such as my formulation of water flow in liquid and vapor phases in plants on a common footing, deficit in mole fraction rather than the formulations used today, water potential in the liquid phase and mole fraction in the vapor phase.
- Here’s some research that recently (Dec. 2017) “leaked” into teaching, as well – a lot about light scattering and absorption
- Biological modeling. I’ve done this extensively, almost exclusively on plants and their exchanges of mass, energy, and momentum with the environment. I’ve seen my share of good and bad models, so that I’ve developed a deep philosophy of biological modeling. This may prove of use in evaluating models or planning efforts in modeling. In a particular topic, optimizatoin modeling, Tom Buckley, now of the University of Syndey, and I have converged on a strong aversion to the common application of optimization.